The CAR is a chimeric transmembrane receptor with an extra-membrane domain, responsible for antigen CD19 recognition via a single-chain variable fragment structure. It is linked through a transmembrane domain to an intracellular component responsible for the T cell activation against its target. Commercial CAR-T cells are obtained from autologous lymphocytes passing through a positive selection of T cells and viral transduction of the CAR. During the manufacturing process, lymphocytes are stimulated with a culture medium containing interleukin 2. Such an expansion process aims at obtaining a target dose that varies according to the commercial product. Second-generation CAR-T cells activation is mediated by co-stimulation of CD-3zeta. Despite the deepening knowledge of CAR-T cells expansion kinetics and resistance mechanisms, few data are available concerning morphological features of these artificially modified cells in ex vivo samples before infusion.
We collected samples of CAR-T cells from the leftovers of infusion bags of two patients with refractory aggressive non-Hodgkin B cell lymphoma, treated at our center with commercial axicabtagene ciloleucel CAR-T cells. After the complete infusion, residual CAR-T cells were obtained from the infusion bags by injection and washing with five milliliters of saline solution. The recovered material was then divided into three aliquots for the analyses. First, a nucleated cell count was performed on an ADVIA® 120 automated cytometer. Second, cells were analyzed by flow cytometry according to the method described by Magnani et al.[1] Data were acquired on BD FACSCantoII flow cytometer and analyzed by BD FACSDiva software (BD Biosciences). Finally, a third aliquot was used to prepare cytospins (800 rpm for 3 min). Peripheral blood samples and smears of treated patients were similarly analyzed during a two-week follow-up. Smears and cytospins were stained according to May-Grümwald-Giemsa and observed at the optical microscope.
The nucleated cell counts of the residual samples were 0.5 and 0.6 x109/L, respectively. Therefore, the analyzer classified all the cells as either lymphocytes or large unstained (myeloperoxidase-negative) cells (LUCs).
At the microscopic examination of cytospin preparations, the lymphoid cells appeared morphologically heterogeneous. We identified the following subtypes: i) large granular lymphocytes (LGL), often with prominent granulations (less than 5%); ii) small-to-medium-sized lymphocytes (30-40%), with moderately abundant basophilic cytoplasm, thickened and hyperchromatic chromatin, and pleiomorphism of the nucleus profile, accentuated indeed by the cytospin preparation; nuclear indentation was however sometimes visible in cells from the peripheral blood smears (Figure 1C); iii) large lymphocytes (25-30%) with promonocytoid features (nuclear indentation, immature chromatin, grey cytoplasm with fine scanty granules); iv) giant/enormous lymphoid cells with blastic features (25-30%), immature chromatin, multiple molded nucleoli, often with reinforced chromatin border, abundant immunoblast-like hyperbasophilic cytoplasm, with numerous purple granulations, with frequent vacuoles and a hypertrophic clear para-nuclear Golgi zone (Figure 1). Apoptotic and smudge cells were also present and not considered in the cell counts.
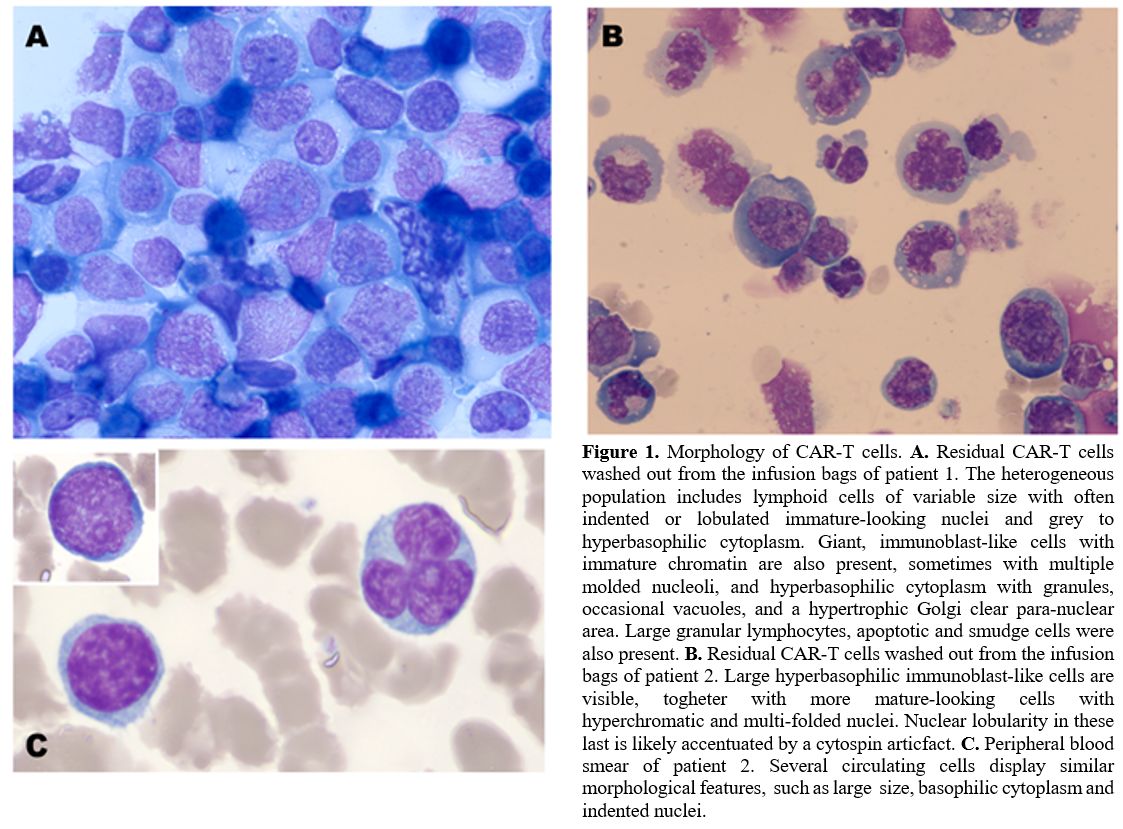 |
Figure 1. Morphology of CAR-T cells. A. Residual CAR-T cells washed out from the infusion bags of patient 1. The heterogeneous population includes lymphoid cells of variable size with often indented or lobulated immature-looking nuclei and grey to hyperbasophilic cytoplasm. Giant, immunoblast-like cells with immature chromatin are also present, sometimes with multiple molded nucleoli, and hyperbasophilic cytoplasm with granules, occasional vacuoles, and a hypertrophic Golgi clear para-nuclear area. Large granular lymphocytes, apoptotic and smudge cells were also present. B. Residual CAR-T cells washed out from the infusion bags of patient 2. Large hyperbasophilic immunoblast-like cells are visible, togheter with more mature-looking cells with hyperchromatic and multi-folded nuclei. Nuclear lobularity in these last is likely accentuated by a cytospin articfact. C. Peripheral blood smear of patient 2. Several circulating cells display similar morphological features, such as large size, basophilic cytoplasm and indented nuclei. |
The immunophenotype evaluation of the two residual samples showed that 76% and 70% of cells, respectively, were CD3+ CAR-T cells. Of these 59% and 63%, respectively, were CD8+ and 39% and 35%, respectively, were CD4+. The median values of side scatter (SSC) and forward scatter (FSC) were increased in both the CD4+ and the CD8+ CAR-T compared to non-CAR-T cells (Figure 2). The largest cells with the highest FSC values mostly consisted of CD4+ CAR-T cells. Their morphology is consistent with giant hyperbasophilic cells with hypertrophic Golgi clear para-nuclear area, as described in population iv.
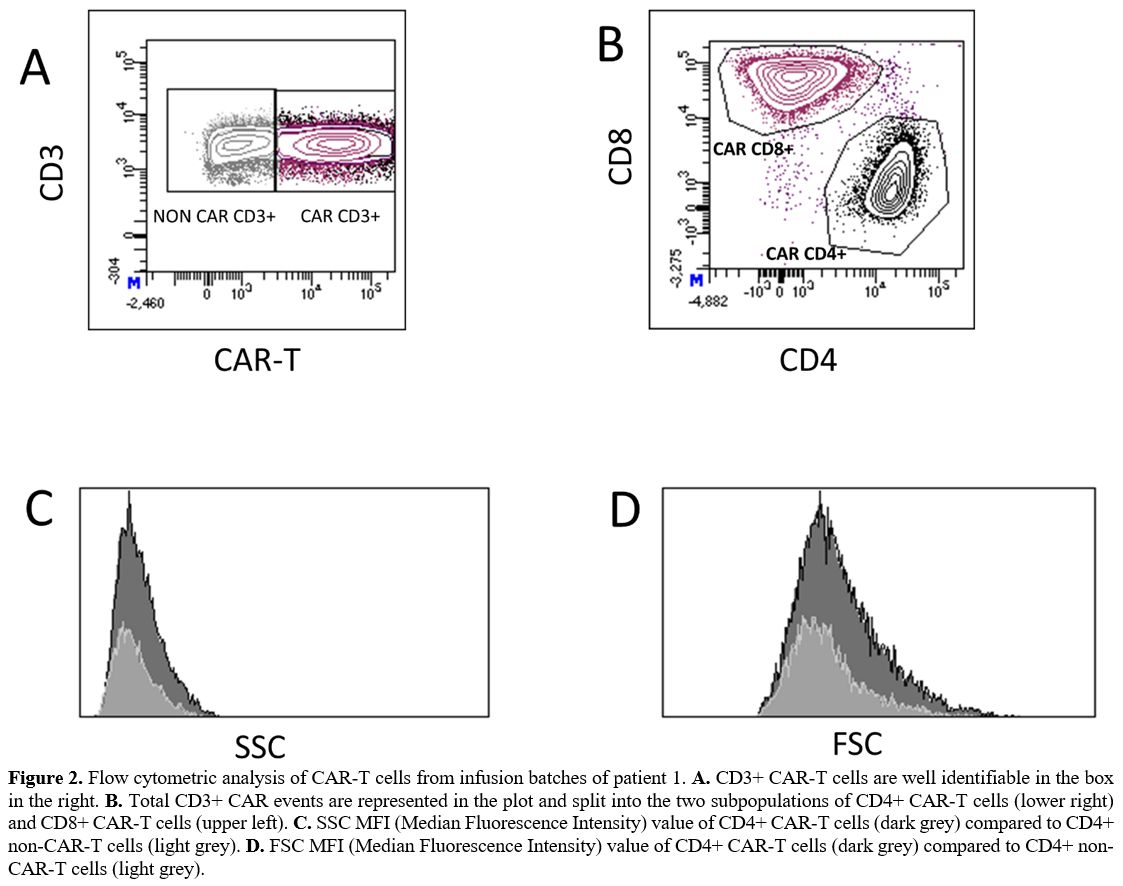 |
Figure 2. Flow cytometric analysis of CAR-T cells from infusion batches of patient 1. A. CD3+ CAR-T cells are well identifiable in the box in the right. B. Total CD3+ CAR events are represented in the plot and split into the two subpopulations of CD4+ CAR-T cells (lower right) and CD8+ CAR-T cells (upper left). C. SSC MFI (Median Fluorescence Intensity) value of CD4+ CAR-T cells (dark grey) compared to CD4+ non-CAR-T cells (light grey). D. FSC MFI (Median Fluorescence Intensity) value of CD4+ CAR-T cells (dark grey) compared to CD4+ non-CAR-T cells (light grey). |
In conclusion, here we describe CAR-T cells' morphological and flow cytometry characteristics from the cell product of two patients at the time of infusion. We identified, among others, a peculiar population of giant lymphoid cells with blastoid features and hypertrophic Golgi clear para-nuclear area. As we identified these cells as CD4+ CAR-T cells, these morphological features most likely reflect massive helper T-cell activation. This activation involves microtubules and the actomyosin cytoskeleton leading to an expansion of the microtubule-organizing center (MTOC).[2,3] Notably, similar large or giant cells, sometimes with nuclear indentations, were also visible with very low frequency on peripheral blood smears ten days after the infusion, consistent with flow cytometric detection of rare CAR-T cells on the same PB samples. Similar cell images have also been reported in bone marrow or peripheral blood smears.[4–6]
Our direct observation of lymphoid subsets with peculiar morphological characteristics in the infusion bag provides a description of morphological features of CAR-T cells and a proof of concept that these are not acquired in vivo secondary to the encounter with the antigen but already present at the time of infusion secondary to viral transduction and in-vitro stimulation. Further studies will be critical to verify if different activation protocols and co-stimulatory molecules in CAR construct, period of resting before sample freezing, and other preparation factors have a role in determining the heterogeneous morphology of these cells.
From a practical standpoint, we underline that the observation of apparently abnormal or immature cells in peripheral blood smears obtained after CAR-T cell infusions should not be immediately be ascribed to the possible occurrence or relapse of a malignant disorder.